
By Dr. Jordi Gómez i Prat and Dr. Sheila MF Mendonça de Souza / Jan. 2003
Prat: Unitat de Malalties Tropicals, Importades i Vacunacions Internationals, D.A P. Ciutat Vella, Barcelona, Catalunya, España
de Souza: Departamento de Endemias Samuel Pessoa, Escola Nacional de Saúde Pública, Fiocruz, Rio de Janeiro, RJ, Brasil

Abstract
Tuberculosis is a prehistoric American human disease. This paper reviews the literature and discusses hypotheses for origins and epidemiological patterns of prehistoric tuberculosis. From the last decades, 24 papers about prehistoric tuberculosis were published and 133 cases were reviewed. In South America most are isolated case studies, contrary to North America where more skeletal series were analyzed. Disease was usually located at the deserts of Chile and Peru, Central Plains in USA, and Lake Ontario in Canada. Skeletal remains represent most of the cases, but 16 mummies have also been described. Thirty individuals had lung disease, 19 of them diagnosed by the ribs. More then 100 individuals had osseous tuberculosis and 26 also had it in other organs. As today, transmission of the infection and establishment of the disease were favored by cultural and life-style changes such as sedentarization, crowding, undernutrition, use of dark and insulated houses, and by the frequency of interpersonal contacts. The papers confirm that despite previous perceptions, tuberculosis seems to have occured in America for millennia. It only had epidemiological expression when special conditions favored its expansion. Occurring as epidemic bursts or low endemic disease, it had differential impact on groups or social segments in America for at least two millennia.
Emergence and Origins of Tuberculosis in America
The origins of human tuberculosis are not yet completely understood. The most traditional hypothesis proposes that the contact with Mycobacterium bovis occurred when the aurochs (Bos primigenus) were domesticated, about 8,000 years ago (Nowak 1991). The direct and frequent contact with the animals and the use of contaminated milk and beef were probably responsible for the introduction of tuberculosis among the human groups. According to Cockburn (1967) crowding and confining animals with humans improved the chance of transmission and illness. The first sedentary settlements that evolved to urban settlements probably favored the selection of the respiratory human form of tuberculosis caused by M. tuberculosis. Changes in lifestyle and oscillation in general conditions of health explain in the past, as today, the different epidemiologic behaviours of tuberculosis in different times and places (Cockburn 1967).
The traditional model also proposes that the small nomadic groups would not fullfil the conditions to keep lung tuberculosis as a disease in the population, contrary to what is expected in bigger, sedentary groups. Otherwise, considering that tuberculosis is a chronic, long-duration disease and Mycobacterium can assume long resistant forms, inside or outside the body, some authors like Johnston (1995) propose the persistence of tuberculosis at low endemicity levels even in the small populations. It would be possible that pulmonary human tuberculosis occurred in human populations even before the Neolithic revolution, in Europe and America as well.
Finally, considering the existence of associations between M. bovis and M. tuberculosis in the clinical cases, and the variations in clinical behaviour of the infection caused by different mycobacteria (Gimenez et al. 1993) the explanations about the evolution of tuberculosis and its emergence in America is under continuos review (Buikstra 1981, Paulsen 1987). With regard to more ancient cases of lung tuberculosis discovered in America, bacterial epidemiology adds to the debate and may result in the proposal of new explanations. Either the ancient prehistoric hunters brought the pathogens from the Asian/European stocks and kept the infection at low endemic levels, or the pathogens were acquired directly from environmental reservoirs in America. Of course, other migrations bringing the disease could certainly be considered, since the archaeologists have proven the existence of multiple migrations.
The terrible bursts of tuberculosis among American Indians after the intensification of contacts with Europeans and some immunologal research points to low immunity among the natives, reinforcing the interpretation that American Indians were “virgin soil” for the infection. Even though this is not completely wrong, data are not sufficient to support the concept that tuberculosis was absent in America previous to the contact. Many paleopatho-logical investigations in the last three decades confirmed the presence of the disease in different prehistoric ancient American populations, and more complex explanations are required. Factors like different pathogenic strains of the bacteria, the absence of long or permanent immunity and, of course, the socio-cultural determinants for the emergence of the disease have to be carefully considered.

Fig. 1: distribution of confirmed cases of prehistoric tuberculosis in North and South America – NA1. Pueblo Bonito. NA2. Pennsylvania. NA3. Memphis. NA4. North Arizona. NA5. West Central Illinois. NA6-1. Hamilton. NA6-2. Williamson. NA7-1. Mobridge. NA7-2. Larson. NA8. Glen Williams. NA9. Saskatchewan. NA10. Ontario. NA11. Uxbridge. NA12. North Dakota. NA13. Moundville. NA14. Irene Mound. SA1. Cusco. SA3. Nasca. SA4-1. Chongos. SA4-2. Montegrande. SA4-3. Huayuri. SA4-4. Lampilla. SA4-5. Murga. SA4-6. Caserones. SA4-7. Caserones. SA4-8. Caserones. SA4-9. Pica. SA4-10. Azapa. SA4-11. Arica. SA5. Estuquiña. SA6. Nasca. SA7. Arica. (Draw: J Prat) / Click Image to Enlarge
Many prehistoric American Indians are described as healthy and not suffering from tuberculosis at the time of the contact, but the primary documents from the XV and XVI centuries generally lack medical details (Paulsen 1987). The absence of archaeological information about them make it difficult to know wether tuberculosis existed or not. In some contact period descriptions of diseases, even when supposed lung tuberculosis is reported, it is impossible to distinguish pre-existent from acquired disease. For people like the Inca, there is paleopathological evidence to prove that they had already been exposed to the infection in their homeland for millennia, and that the infection was possibly endemic when the European arrived. It is likely that different susceptibilities existed, and different selective and imunological conditions existed, as a result of different epidemiological histories in America. In all cases, social and cultural costs of contact caused a big health impact (Verano & Ubelaker 1992) and tuberculosis was probably one of the most sensitive indicators of those historical and biocultural processes.
To discuss the entrance of tuberculosis in America it is also necessary to understand the entrance of humans in America, another debated subject. Models currently accepted consider that non-mongoloid and mongoloid human groups of Homo sapiens entered America in subsequent waves, at least during the last 20,000 to 15,000 years ago (Salzano & Callegary-Jacques 1988, Neves et al. 1997). The last successful prehistoric migration came along the Arctic calotte about 6,000 years ago, bringing the Eskimo ancestors. The only known contact between the post-Neolithic Europeans and American Indians is the unsuccessful Viking attempt to colonize North America during the XI century. Regular contacts between Old and New World started after the XV century, and still tuberculosis is far more ancient in North and South America, according to archaeological data. The logical interpretation is that tuberculosis has emerged in America in an independent way.
Clark et al. (1987) proposed four main possible explanations for the existence of prehistoric tuberculosis in America: 1. Was caused by the exposure of human populations previously immunized by free Mycobacterium to the M. tuberculosis, 2. Was caused by less virulent variations of M. tuberculosis, and that would also explain the high susceptibility of modern Indians to tuberculosis, 3. Was caused by M. bovis acquired from wild fauna like the bison, and this would also explain the bone lesions, 4. Was caused by different varieties of free Mycobacterium, and this also would explain the high susceptibility to the European varieties of bacteria. Changes in the knowledge about tuberculosis, like the concept of “tuberculous complex”, and the recent paleoepidemiologic data about the respiratory form of the disease in the prehistory add information to a necessary review of previous models.
The oldest North American description of prehistoric tuberculosis is from Tennessee (Putman & Whitney 1886 in Morse 1961). Modern methodologies applied to that material and others, confirmed positive results (Morse 1961). Many papers were published about paleopathology of tuberculosis, but most of them offer gross descriptions and have no safe chronology. Thus many cases of tuberculosis are of either doubtfull diagnosis, or could be post-contact, so that it is not possible to consider them. Anyway, two decades after the printing of the most important review for tuberculosis in America (Buikstra 1981), new data has been added to the literature especially about South America. Also tuberculosis remergered as a more a frightening health problem all over the world. Because of these new factors, it is time to go back to the theme of the origins of tuberculosis in the New World.
The present revision and update of the literature reviews 24 papers of the last decades that describe 133 cases of prehistoric tuberculosis. South American cases include isolated descriptions. The main locations of the disease there are the deserts of Chile and Peru. North American cases, mostly represented by skeletal series, were found at the Central Plains and Lake Ontario, in USA and Canada. Tuberculosis is more represented by skeletal lesions but could also be diagnosed at other anatomical sites in 16 mummified bodies. Confirmation of the importance of the respiratory form of the disease is based on the description of 30 cases of lung tuberculosis.
South America: Prehistoric Findings
The first undoubted confirmation of tuberculosis in South America was published by Allison et al. (1973) who described a mummified child of 8 to 10 years from Hacienda Agua Salada, a site close to Nasca, Peru. Associating pathological analysis of mummified remains, radiology and bacteriological studies, the diagnosis was confirmed subsequently by other techniques in five different publications. After that successful microscopic demonstration of mycobateria in archaeological material (Allison et al. 1981) other pre-Columbian samples were checked and other cases of tuberculosis could be diagnosed in eleven different mummies from Chile and Peru. Years later Buikstra and Williams (1991) found 37 cases of tuberculosis in Southern Peru, publishing the first paleoepidemio-logical study for this infection in South American collections. It was confirmed by Salo et al. (1994), who published the first molecular diagnosis using PCR tecniques for mycobacteria’s aDNA in the same material. A second molecular diagnosis was published by Arriaza et al. (1995), for one Arica mummy. Most of the available publications for South America are case studies and do not really contribute to paleoepidemiologic approaches, but Buikstra and Williams (1991) and Arriaza et al. (1995) could analyse skeletal series, either contemporaneous or from different periods, proposing the first population approach to the disease (Table I).

Table 1: Click Image to Enlarge
South American descriptions of tuberculosis represent a big contribution to the understanding of its prehistory because they provide information about well-preserved soft tissues such as lungs. This is especially important for proving the existence of respiratory forms of the disease. Other important contributions of South American descriptions include the involvement of different organs, the demonstration of the milliary form of the disease, and the similarity between ancient tuberculosis and the modern clinical/pathological form.
The present spacial distribution of tuberculosis in South America forces us to consider the preservation bias because most of the findings were in desert areas where mummies and skeletons are well preserved. Mummified bodies found at Southwestern Andes and even Cuzco, 3,300 m above the sea, also had tuberculosis. Only one case was described outside of Chile and Peru, in El Palito, Middle West of Venezuela’s coast .

Fig. 2: Wari archaeological urban settlement in Peru (Photo: J Prat)
Tuberculosis existed in South America for almost 2,000 years. The most ancient case belongs to the Paracas-Caverna culture, dated around AD 160. Subsequently, other cases were found at Hacienda Água Salada and Cahuachi, Nazca sites around AD 200 to 700. The next cultural period, of the Wari, lasted until AD 1100 and tuberculosis was also found among their human remains, at Montegrande and Huayari. A burial site of the Chiribaya, culture that followed the Wari, was escavated at Estuquina, Peru and 37 cases of tuberculosis were confirmed. Chiribaya belongs to the so called Formative period that preceeded the Regional States, during this period social complexity was developing. The last prehistoric findings, close to the contact period, belong to the Inca culture. Considering that individuals belonging to the subsequent and different cultures at the same region had the respiratory and/or bone infection, it is possible to propose that tuberculosis persisted as an endemic health problem there for two millennia, and was still there at the XVI century when the Europeans arrived.
Whether case studies or population studies, the results from South America help to confirm the existence of tuberculosis at least in Atacama Desert and Andean proximities for 2000 years of continuous human occupation. Affecting the main prehistoric groups in the region, tuberculosis maintained a transcultural persistent pattern, from the Formative period to the European contact. Cultural interactions and common biocultural background certainly explain the persistence and periodic expansion of this disease in South America. People like the Paracas, who created villages of dark insulated pithouses (Caceres Macedo 1999) probably maximised their risk of infection and disease. The occurrence of sporadic cases or low endemicity tuberculosis probably increased in periods such as the Paracas and therefore increased the chance of finding tuberculosis in archaeological remains. The Wari were characterized by the urban process and building of big fortified cities (Cáceres Macedo 1999). In these cities, tuberculosis (Allison et al. 1981) was probably favoured by crowding and interpersonal contacts inside the urban walls. The Inca, with their small, dark, badly ventilated houses of only one room, 5 to 6 m large, certainly sustained tuberculosis (Garcia-Frias 1940, Allison et al. 1981, Soriano 1997), and their well organized system of communications probably helped to disseminate the disease among different people living in Tawantinsuyo, under the powerfull network of their empire.
Paleoepidemiologic investigations suggest that tuberculosis could also have occurred as epidemic bursts. In Estuquina, Peru (Buikstra & Williams 1991) the protective wall built around the Chiribaya city, sequestered the population in times of war, and must have favoured both transmission and disease. In Arica, Chile, a similar process is proposed by Arriaza et al. (1995) to explain his findings. Changed lifestyle from living in small spread groups during the Cabuza-Tiwanaku phase, to living in big urban settlements during the Maitas-Chiribaya phase, was probably a disruption factor causing the burst of the disease. Different epidemiological behaviours probably also occurred in the past. In Estuquina there are many infections in young adults, 20 to 29 years old, and in older adults, more than 50 years old, but only a few children show signs of tuberculosis. This suggests they probably were a previously exposed population, and that illness resulted from temporary frailty. In the same cemetery, men are three times more affected then women, suggesting a specific risk for males, perhaps stressed and crowded because of their role in wars. Other population studies will certainly help to propose comparative models.
The varied anatomical distributions of the lesions in mummies prove tuberculosis was not different from the present infection. Infected lymphnodes, lungs, bones, meninges, kidneys, and also miliary tuberculosis were found (Table II). Comparing soft tissue lesions and rib periosteal reactions with bone tuberculosis for the same individuals in South American collections will be the first step to confirm the application of epidemiologic models to the interpretation of paleopatholological findings.
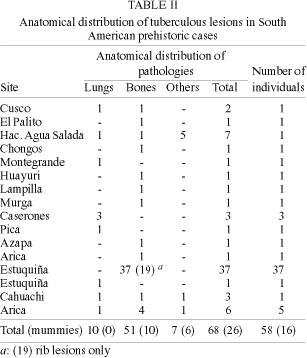
Table 2: Click Image to Enlarge
North America: Prehistoric Findings
Eighteen papers published in the last decades about North America prehistoric tuberculosis are more uniform than South American literature regarding materials and methods, as well as the results.
Judd (1954) described the first dated case for tuberculosis in North America at Pueblo Bonito, an Anasazi site, circa AD 828 to AD 1130. This case was revised by El-Najjar (1979) some years later together with two other cases from Chavez Pass, northern Arizona dated AD 900 to AD 1100. Three papers published in 1985 documented the presence of tuberculosis in several pueblo time periods and locations. Sumner (1985) found a case of possible tuberculosis in an early pueblo village in northeastern Arizona and demonstrated that this disease was present among the earliest puebloans. Fink (1985) documented a case of a child infected with tuberculosis from the Chuska Mountains on the Arizona, New Mexico border. The child died at the height of Pueblo cultural development in the region and demonstrates that the disease was endemic. Micozzi and Kelley (1985) found tuberculosis in the Point of Pines Site in the White Mountains of Arizona. This is a late, Eastern Pueblo site. Thus, tuberculosis was a health problem for Pueblo people at all times. Fink (1985) describes the epidemiological conditions that would have promoted the disease in the Pueblos. These conditions include lack of knowledge of the germ theory, crowded living conditions, communal eating utensils, poor ventilation, and poor exposure to sunlight.
The more important contribution to the theme was the study published by Buikstra and Cook (1981) who reviewed skeletons from different sites of the Mississippi culture, in West Central Illinois, representing a long cultural sequence from 150 BC to AD 1150. In this study, a chronological sequence of prevalences for different periods allowed the authors to associate the emergence of the infection to cultural changes. In another paper Widmer and Perzigan (1981) describe seven cases of tuberculosis in two other sites, one from the Mississippi culture, in Ohio, dated from AD 1275; and another from Middle Cumberland culture, in Tennessee, dated from AD 1200. Palkovich (1981) published a prehistoric case of tuberculosis from an Arikara site in South Dakota. In Canada, Hartney (1981), Walker (1983) and Melbye (1983) described tuberculosis in Ontario. Pfeiffer (1984) found 18 cases of tuberculosis in Uxbridge, another Iroqui site in Ontario. Four other cases are described by William and Snorthland-Coles (1986) in Jamestown Mounds, USA, and by Powell (1988) in Moundville, a Mississipi culture site in Alabama, and in Irene Mound, a site in Georgia (Table III).

Table 3: Click Image to Enlarge
Contrary to what happened in South America, more population studies were done in North America (Buikstra & Cook 1981, Palkovich 1981, Hartney 1981, Pfeiffer 1984, William & Snortland-Coles 1986, Powell 1988) and more consistent paleoepidemiologic discussions about tuberculosis were possible. The contribution of Buikstra and Cook (1981) was especially important because besides considering the specific signs for diagnosis in bones they also considered the prevalence of unspecified stress as indicators of infection.
Contrary to the spacial distribution in South America, probably biased by preservation, in North America a great number of sites have been excavated and many skeletal remains studied. Perhaps its distribution in Central Plains, especially along the Mississippi River basin is more close to the real geographic distribution of the prehistoric disease on the continent. Middle Woodland culture (AD 150-400), Late Woodland (AD 400-1050) and Mississippi (AD 1050-1150) culture have furnished good information about tuberculosis and its association to socio-economic changes (Buikstra & Cook 1981, Buikstra 1981). The increasing population, a subsistence strategy based in exchange of surplus products, periodical meetings in the ceremonial and commercial centres, and bad nutrition are some of the conditions that probably favoured the increasing number of cases in the Mississippi basin. According to McGrath (1987) there were differences in the settlements in the three periods: from independent small-scattered groups they evolved to bigger and interdependent groups, meeting periodically in Cahokia, an urban economic centre that probably had 10,000 to 30,000 inhabitants. The hypothesis of endemic tuberculosis in such big villages can be easly supported by any epidemiologic model.
In the Mississippi basin, as in other places, the recurrent interpersonal and inter-group contacts also seem to have had importance in the dispersion of tuberculosis. This factor can explain the presence of the infection in different neighbor cultures, despite their different life-styles (Lobato & Hopewell 1998), since population mobility is an important risk factor for tuberculosis (Casper et al. 1996, Hermans et al. 1995, Yang et al. 1994, Bifani et al. 1996). Williams and Snortland-Coles (1986) described tuberculosis in nomadic groups that regularly exchanged goods with big sedentary groups in Missouri, proposing that the contacts increased their risk of infection. A model like this would well explain prehistoric tuberculosis crossing America with the help of cultural interaction spheres, reaching also the nomadic peoples. In other regions like the Sowthwest, local factors seem to have contributed to the emergence of the disease like those proposed by Fink (1985) and Reinhard et al. (1988) for Anasazi Pueblos where aerial transmission of infections probably followed architectural and economic changes.
In North America, contrary to what happened in South America, none of the cases were confirmed by aDNA, bacteriology, or soft tissue examinations. All the cases are represented by bone infection. Only one skeleton had adherent pleural tissue confirming pleural lesion. Most of the 72 cases were diagnosed in adults of different ages (Table IV). A possible bias, considering the nature of funerary samples (Waldron 1994) is suggested by the fact that only Buikstra and Cook (1981), using a paleoepide-miologic approach, were able to propose the inclusion of children less than 12 years in their ill series. As sex determination of badly preserved skeletons is dubious, only in Turpin Mound and Irene Mound collections is a possible sex ratio discussed. Tuberculosis seems to be more prevalent in women from Turpin Mound (2:1 and 3:1 respectively) and in men from Chavez Pass (3:1). Although the age distribution of cases is not clear, the prevalence differences for men and women can be suggestive of changing epidemiologic patterns according to biocultural specificities.
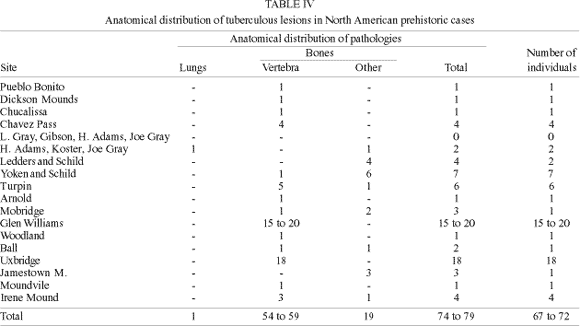
Table 4: Click Image to Enlarge
New Paleoepidemiologic Interpretations?
For a long time it was the consensus that the Europeans brought tuberculosis to America. Archaeological data succeeded in proving that the disease was already in America at least 2,000 years before the contact. More than just existing, tuberculosis had already assumed an epidemiologic pattern of aerial transmission. It was initially a sporadic infection but later became a very significant health problem for some groups. On the other side, literature review also shows that in spite of the advanced techniques available today, the paleopathological information is still insufficient for a complete comprehensive epidemiological reconstruction of tuberculosis in America.
Some of the models proposed in the last decades to explain the origins of prehistoric tuberculosis (Cockburn 1967, Clark et al. 1987) do not fit American data any more. Enteric sporadic tuberculosis, or opportunistic infection caused by free mycobacteria, cannot be the only explanation considering the number of prehistoric cases, and the temporal persistence and spacial distribution of those cases. Paleoepidemiological studies in North America, and the study of mummy and skeletal remains in South America, prove the existence of lung disease that progressivelly acquired more epidemiologic importance as an air borne disease. Tuberculosis in the past is strongly associated to sedentary life-styles, certain special patterns of architecture and urbanisation, as well as to the existence of social disrupting factors, as it is today.
In many places, tuberculosis probably persisted in low endemic patterns and populations could be protected by the so-called herd immunity phenomenon. The same explanation could be applied in the case of some Indian groups, if life conditions before and after contact were compared. Social disruptions and other biocultural factors certainly elevated the number of susceptibles to mycobaterial infections.
Considering this fact will certainly help to develop a hypothesis that can help in searching for other archaeological sites where tuberculosis could be present in the past. Considering the state of art, the absence of tuberculosis in many parts of America, especially in South America, must still be considered absence of information, instead absence of infection.
Along the last 2000 years of subsequent different cultural periods, in both Americas, there is archaeological evidence of tuberculosis. The dispersion and persistence of the disease strongly suggest the antiquity of the tuberculous complex that probably came to America with the first human migrations. Among the many species of fauna that could be reservoirs to mycobacteria a few examples are the buffalo and the wild cats and dogs, that can be infected by M. bovis and M. tuberculosis, and birds like the turkey, that keep the M. avium (Hubert 1975). Free mycobacteria, some of them found in the water springs, could also have participated of the epidemiological scenario (Stead 2000). The transition from the hunt-ering-gathering subsistence pattern to the sedentarization brought many changes in life pattern, including partly-domesticated animals like the camelids (Llama sp.) and the guinea pig (Cavia sp.) living in intimacy with humans, besides other occasionally hosted in settlements, like the wild dogs. Although they are not natural hosts for the main bacteria of the tuberculous complex, they could occasionally have shared the human infection.
As soon as Allison et al. (1973) proved the existence of pulmonary tuberculosis in archaeological material, the following researchers were able to demonstrate its importance to the epidemiological scenario of prehistoric America. But the rarity of mummified material limited the possibilities of investigation, as demonstrated by the North American papers. More recently, the study of pleural rib periostitis provided additional models to test lung tuberculosis, i.e. air borne transmission, for even in skeletal remains (Kelly & Micozzi 1984, Santos 2000). From 16 cases of tuberculosis in Peruvian mummies, 10 (62.5%) are lung diseases (Garcia-Frias 1940, Allison et al. 1973, Allison et al. 1981, Salo et al. 1994, Almonacin 1994, unpublished results, Arriaza et al. 1995). From 37 cases of tuberculosis found in Estuquina, 19 (50.1%) have rib periostitis (Buikstra & Williams 1991). At least in South America, respiratory tuberculosis was certainly an important health problem. Recent investigations in skeletal samples from Solcor 3 and Quitor 6, sites from São Pedro de Atacama, Chile, strongly suggested tuberculosis as a cause of death (Mendonça de Souza & Gómez i Prat 1999).
Special conditions in South America like the Nazca wars, the Wari commercial activities (Cáceres Macedo 1999), the transhumance pattern of mobility and the cultural interaction spheres that molded the rich net of contacts between peoples in highlands and in lowlands (Nuñez & Dillehay 1995) probably favoured the expansion of the disease. On the other hand, the Moche were apparently more isolated and could have had less infection.
If tuberculosis was in America, especially in the urban centers and culturally sophisticated states, it is possible to propose that the European arrival suddenly disrupted the epidemiological scenario. The contact certainly made tuberculosis more potent in many different ways by bringing new varieties of mycobacteria, especially to areas where the disease was rare, decimating Indians with the help of social disruption, adding new viral infections as measles, moving people, and just imposing stress. In any case, tuberculosis came to be one of the American Indian plagues, as even the Inuit of the XX century can easily testify. Could the American Indians have an alternative history with tuberculosis without the European contact?
Tuberculosis is a disease of poverty and social disruption, whether pre-existing or introduced. It bursts, today as yesterday in impoverished conditions. In North and South America prehistoric tuberculosis followed where quality of life declined. Although many authors disagree about the use of the opposition “nature x nurture” in tuberculosis, advising about the danger of reducing the disease to its social predisponent factors (Stead 2000), it is not a coincidence that tuberculosis in the prehistoric past, as in the present, was an opportunistic disease growing together with bad health, nutrition, crisis, war, ignorance, crowding, and confinement.
An intriguing point in paleoepidemiology of tuberculosis in prehistoric North America is that most of the cases of tuberculosis are dated from the second millenium, after the Vikings with their domesticated cattle settled in colonies on the Eastern Coast. According to written documents they exchanged goods, including cow milk, with Indian groups, and this fact forces us to consider the possible penetration of European strains of mycobacteria in North America previous to the XV century. Perhaps North and South America have different epidemiological histories for tuberculosis, and more aDNA studies are necessary to define the varieties of pathogenic bacteria in different regions, the prevalence of enteric tuberculosis in South America, and other details that will certainly help to draw paleoepidemiologic scenario in both continents.
The absence of data for Amazon and Mesoamerica, where the preservation is poor because of the climatic conditions, is an important barrier to learning more about tuberculosis past geography. In Amazonia, as in some of the archaoelogical regions described here, big human groups of thousands of people in dense populations could have sheltered prehistoric tuberculosis. Mesoamerica urban centers certainly offered many of the conditions described in the literature for North and South America. The biocultural conditions that could have promoted the development of the disease in some of those places were as adequate as in Cahokia or Estuquina, and perhaps Amazonian or Mesoamerican peoples also shared periods of expansion of the disease.
Paleoepidemiologic studies are very difficult because funeral series are seldom good for proper analysis and inference. Prehistoric tuberculosis is a very interesting subject but only the last two decades of intensive studies brought us data of epidemiologic value, and after the recent repatriation acts in the United States the chance of studying the North American archaeological series is strongly reduced. Perhaps only in South America can future archaeological and paleopathological research shed light on the past epidemiology of tuberculosis through its extraordinary sites and collections.
References
Allison M, Gerszten E, Munizaga J, Santoro C, Mendoza D 1981. Tuberculosis in pre-Columbian andean populations. In JE Buikstra, Prehistoric Tuberculosis in the Americas, Northwestern University Archaeological Program, Evanston, p. 49-61.
Allison M, Mendoza D, Pezzia M 1973. Documentation of a case of tuberculosis of pre-Columbian America. Am Rev Resp Dis 107: 985-991.
Almonacin GL 1994. Deteccion de Mycobacterium tuberculosis en uma Momia de la Cultura Nasca com Mal de Pott, Facultad de Medicina Alberto Hurtado, Lima, 59 pp. (mimeo).
Arriaza BT, Salo WL, Aufderheide AC, Holcomb TA 1995. Pre-Columbian tuberculosis in Northern Chile: molecular and skeletal evidence. Am J Phys Anthropolol 98: 37-45.
Bifani PJ, Plikaytis BB, Kapur V, Stockbauer K, Pan X, Lutfey ML, Moghaseh SL, Eisner W, Daniel TM, Kaplan MH, Crawford JT, Musser JM, Kreiswirth BN 1996. Origin and interstate spread of a New York city multidrug-resistant Mycobacterium tuberculosis clone family. J Am Med Assoc 275: 452-457.
Buikstra JE 1981. Prehistoric Tuberculosis in the Americas. Northwestern University Archaelogical Program, Evanston, 182 pp.
Buikstra JE, Cook DC 1981. Pre-Columbian tuberculosis in West-Central Illinois: prehistoric disease in biocultural perspective. In JE Buikstra, Prehistoric Tuberculosis in the Americas, Northwestern University Archaelogical Program, Evanston, p.115-139.
Buikstra JE, Williams S 1991. Tuberculosis in the Americas. In DJ Ortner, AC Aufderheide (eds), Human Paleopathology. Current Synthesis and Future Options, Smithsonian Institution Press, Washington, p.161-172.
Cáceres Macedo J 1999. Las Culturas Prehispánicas del Perú. Guía de Arqueologia Peruana, Lima, p. 61-71, 125-134.
Casper C, Singh SP, Rane S, Raley CH, Schecter GS, Riley LW, Kreiswirth BN, Small PM 1996. Transcontinental transmission of tuberculosis: a molecular epidemiological assessment. Am J Public Health 86: 551-553.
Clark GA, Kelley MA, Granje JM, Hill CM 1987. The evolution of mycobacterial disease in human populations. Curr Anthropol 28: 45-62.
Cockburn A 1967. Infectious Diseases: Their Evolution and Erradication, Charles C Thomas, Springfield, 402 pp.
El-Najjar MY 1979. Human treponematosis and tuberculosis: evidence from the New World. Am J Phys Anthropol 51: 599-618.
Feldman WH 1963. Tuberculosis. In TG Hull, Diseases Transmitted from Animals to Man, Charles C Thomas, Springfield, p. 5-81.
Fink TM 1985. Tuberculosis and anemia in a Pueblo II-III (ca. AD 900-1300) Anasazi child from New Mexico. In CF Merbs, RJ Miller, Health and Disease in the Prehistoric Southwest, Arizona State University, Anthropological Research Papers 34.
Garcia-Frias JE 1940. La tuberculosis en los antiguos peruanos. Actualidad Med Peruana 10: 274-291.
Gimenez Alvez MA, Carvalho Rizzon CF, Picon PD, Milano AC 1993. Tuberculose gastrintestinal. In PD Picon, CFC Rizzon, WP Ott (eds), Tuberculose. Epidemiologia, Diagnóstico e Tratamento em Clínica e Saúde Pública, Médica e Científica Ltda., Rio de Janeiro, p. 405-416.
Hartney PC 1981. Tuberculosis lesions in a prehistoric population sample from southern Ontario. In JE Buikstra, Prehistoric Tuberculosis in the Americas, Northwestern University Archaelogical Program, Evanston, p.141-160.
Hermans PWM, Messadi F, Guebrexabher H, van Soolingen D, Haas PEW, Heersma H, Neeling H, Ayoub A, Portaels F, Frommel D, Zribi M, van Embden JDA 1995. Analysis of the population structure of Mycobacterium tuberculosis in Ethiopia, Tunisia and the Netherlands: usefulness of DNA typing for global tuberculosis epidemiology. J Infect Dis 171: 1504-1513.
Hubert WT 1975. Diseases Transmitted from Animals to Man, Charles C, Thomas, Springfield.
Johnston WD 1995. Tuberculosis. In KF Kiple, The Cambridge World History of Human Disease, Cambridge University Press, New York, p.1059-1068.
Judd NM 1954. The Material Culture of Pueblo Bonito, (Smithsonian miscellaneous collection # 124), Smithsonian Institution, Washington.
Kelley MA, Micozzi M 1984. Rib lesions in chronic pulmonary tuberculosis. Am J Phys Anthropol 65: 381-386.
Lobato MN, Hopewell PC 1998. Mycobacterium tuberculosis infection after travel to or contact with visitors from countries with a high prevalence of tuberculosis. Am J Respir Critic Care Med 158: 1871-1875.
McGrath JW 1987 Social networks of disease spread in the Lower Illinois Valley: a simulation approach. Am J Phys Anthropol 77: 483-496.
Mendonça de Souza SMF, Gómez i Prat J 1999. Paleo-epidemiologia da tuberculose nas Américas. In Resumos da I Bienal de Pesquisas da Fundação Oswaldo Cruz, Rio de Janeiro, p. 407.
Micozzi MS, Kelley MA 1985. Evidence for Pre-Colombian tuberculosis at the Point of Pines Site, Arizona: skeletal pathology in the Sacro-Ilac Region. In CF Merbs, RJ Miller, Health and Disease in the Prehistoric Southwest, Arizona State University, Anthropological Research Papers 34.
Morse D 1961. Prehistoric tuberculosis in America. Am Rev Resp Dis 83: 489-504
Neves WA, Zanini MC, Munford D, Pucciarelli HM 1997. O povoamento da América à luz da morfologia craniana. Rev USP 34: 96-107.
Nowak RM 1991. Walker’s Mammals of the World, John Hopkins University Press, Baltmore, p. 1425-1431.
Nuñez L, Dillehay TS 1995. Movilidad Giratoria, Armonía Social y Desarrollo en los Andes Meridionales: Patrones de Tráfico e Interacción Económica, Universidad Católica del Norte, Antofagasta, 120 pp.
Palkovich AM 1981. Tuberculosis epidemiology in two Arikara skeletal samples: a study of disease impact. In JE Buikstra, Prehistoric Tuberculosis in the Americas, Northwestern University Archaelogical Program, Evanston, p.161-175.
Paulsen HJ 1987. Tuberculosis in the native Americans: indigenous or introduced? Rev Infect Dis 9: 1180-1185.
Pfeiffer A 1984. Paleopathology in an Iroquoian Ossuary, with special reference to tuberculosis. Am J Phys Anthropol 65: 181-189.
Powell ML 1988. Endemic treponematosis and tuberculosis in the prehistoric Southeastern United States: biological costs of chronic endemic disease. In DJ Ortner, AC Aufderheide (eds), Human Paleopathology, Smithsonian Institution Press, Washington, p. 173-180.
Reinhard KJ 1988. Cultural ecology of prehistoric parasitism at Colorado Plateau as evidenced by coprology. Am J Phys Anthropol 77: 355-366.
Requena A 1945. Evidencia de tuberculosis en la América Precolombina. Acta Venezolana 1: 141-164.
Salo WL, Aufderheide AC, Buikstra J, Holcomb TA 1994. Identification of Mycobacterium tuberculosis DNA in a pré-Columbian Peruvian mummy. Proc Nat Acad Sci USA 91: 2091-2096.
Salzano FM, Callegari-Jacques SM 1988. South American Indians: a Case Study in Evolution, Oxford University Press, New York, 242 pp.
Santos AL 2000. A Skeletal Picture of Tuberculosis: Macroscopic, Radiological, Biomolecular and Historical Evidence from the Coimbra Identified Skeletal Collection, PhD Thesis, Departamento de Antropologia, Universidade de Coimbra, Coimbra, 244 pp.
Soriano WE 1997. Los Incas: Economia, Sociedad y Estado en la Era del Tahuantinsuyo, Amaru, Lima, p. 162-166.
Steinbock RT 1976. Paleopathological Diagnosis and Interpretation, Charles C Thomas, Springfield, 423 pp.
Sumner DR 1985. A probable case of Prehistoric tuberculosis from northeastern Arizona. In CF Merbs, RJ Miller, Health and Disease in the Prehistoric Southwest, Arizona State University, Anthropological Research Papers 34.
Verano JW, Ubelaker DH 1992. Disease and Demography in the Americas, Smithsonian Institution Press, Washington, 294 pp.
Walker EG 1983. The Woodland Site: a case for interregional disease transmission in the late prehistoric period. Canadian J Archaeol 7: 49-59.
Widmer L, Perzigan AJ 1981. The ecology and etiology of skeletal lesions in late prehistoric populations from Eastern North America. In JE Buikstra, Prehistoric Tuberculosis in the Americas, Northwestern University Archaelogical Program, Evanston, p. 99-113.
Williams JA, Snorthland-Coles S 1986. Pre-contact tuberculosis in a plains woodland mortuary. Plains Anthropol 114: 249-252.
Yang ZH, Haas PEW, van Soolingen D, van Embden JDA, Andersen AB 1994. Restriction fragment length polymorphism of Mycobacterium tuberculosis strains isolated from Greenland during 1992: evidence of tuberculosis transmission between Greenland and Denmark. J Clin Microbiol 32: 3018-3025.






