

The first forms of recorded expression are thought to be those that co‐evolved with technology some 20,000–30,000 years ago.

By Dr. Luke Strongman
Professor of Library Studies
The Open Polytechnic Institute of New Zealand
Abstract
In the twenty‐first century, there are between 6000 and 8000 different languages spoken in the world, all of which are in a continuous state of evolving, by inter‐mixing or stagnating, growing or contracting. This occurs through changes in the population size of the people who use them, the frequency and form of their use in different media, through migration and through inter‐mixing with other languages. As Stadler et al. argue, human languages are a ‘culturally evolving trait’ and when it occurs language change is both sporadic and robust (faithfully replicated) and the main established variants are replaced by new variants. Only about 200 of these disparate languages are in written as well as spoken form, and most, except the popular ones like Mandarin, Spanish, English, Hindi, Arabic, Portuguese, Bengali, and Russian, are in decline of use. But how did language itself evolve and come to be the most important innate tool possessed by people? The complex issue of language evolution continues to perplex because of its associations with culture, social behaviour and the development of the human mind.
Introduction
There are several opinions as to the origin of human language [1, 2]. These include that language originated in a gesture basis, in primitive vocalisations, or in song; but most probably, a combination of all three contributed to human language development over approximately the last 200,000 years. Language, as much as technology, is the pinnacle of human evolutionary achievement—culture and society would not be possible without language. How is it that novice language learners can acquire a complex knowledge of language without necessarily knowing the grammatical rules and cognitive processes that lie behind it? As Fedurek and Slocome suggest ‘Linguists have been intrigued by this counter‐intuitive gap, known as Plato’s problem, between the quality and quantity of available linguistic data and the complexity of linguistic knowledge from which such inputs are created’ [3]. The nature or nurture debate is perhaps simplistic, but it nevertheless represents in language evolution the convergence of biology with culture. As Jakobson suggested, the question is whether ‘the isomorphism of these two different genetic and verbal codes can be explained by a simple convergence stemming from a similarity of needs, or whether the foundations of the manifest linguistic structures, intimately based on molecular communication, are not directly patterned on the structural properties of the latter’ [4]. Approaches to language acquisition and use have taken one of four paths: first, language structure and how structure is informed by human biology and physiology; second, the ability to acquire language among human and possibly some animal species; third, the functional conditions of language; and fourth, the history of the evolution of language [5].
In contemporary times, most theoreticians agree that language has arisen because of the interaction of three complex adaptive systems: biological evolution, human learning and through the acquisition and sustainment of human culture [6]. However, biological evolution imposes physiological constraints, as does the human environment. Thus, conversely, it may be a poverty of stimulus, which creates the need for a linguistic structure and a genetic and cultural substrate to linguistic development. Humans are physiologically adaptable and intelligent and can fashion their environments after their own conscious thought. Of human capabilities, language is one of the most complex phenomenon and its multi‐valency and ubiquity make it a distinctive property of human culture. As Fedurek and Slocome put it: ‘Language enables humans to represent and communicate complex abstract information, and it occurs in verbal, gestural and written forms’ [7]. Language is also inherently combinatorial, with syntax (sentence structure) leading to semantics (sentence meaning).
The Social Conditions of Language

In order to explain language acquisition, the eminent linguist Noam Chomsky proposed that there is a sophisticated genetically encoded propensity for humans to acquire language, which comprises a Universal Grammar (UG) that undergirds language use, and a language acquisition device (LAD) that guides linguistic competence from observed and learned ‘social’ data [8]. Thus, although there is a physiological substrate for language propensity, the information of language is also located in the wider ‘semiosphere’ negotiated between people and their environment and other people. Language is a shared mental as well as physical (sound and kinetic) construct. It is as if it is a property distributed across communication networks among individual and groups of people, communities and cities. As Starte states, ‘there is no absolute distinction between the evolution of media, language, or that of the human species’ [9]. We cannot conceive of human society without forms of communication and language. It is also often said that ‘language does not fossilise’, by which is meant that language is ephemeral, and linguistic markers are by and large mental constructs that do not indexically substrate in the ecosystem.
The first forms of recorded expression are thought to be those that co‐evolved with technological innovation and took place some 20,000–30,000 years ago. Starte also states that there was a progression from logographic and phonetic writing to forms of syllabic and alphabetic writing [10]. What makes language special is that, although the vocabularies of most language users are limited to 50,000 words or so, those vocabularies are nevertheless constructed in a way that allows for unbounded transmission of messages which, when undergirded by a universal grammar, can result in a high fidelity of communication between language users.
From a bio‐linguistic perspective, Berwick and Chomsky argue that:
we can think of language as, in essence, an ‘organ of the body’, more or less on a par with visual or digestive or immune systems … it is a cognitive organ, like the systems of planning, interpretation, reflection… We can think of language as a mental organ, where the term ‘mental’ simply refers to certain aspects of the world, to be studied in the same way as chemical, optical, electrical and other aspects [11].
The use of language is predicated in humans on the expression of the FOXP2 and CNTNAP2 genes and the population frequency of two brain growth and development genes, ASPM and Microcephalin [12], but the array of possibilities for language use is limited only by cognitive complexity. However, fortunately the human brain at approximately 1350 cc has evolved just for the management of such complexity, and in the human brain, the temporal lobe is 23% larger than other brain regions and four times larger than that of our closest mammalian relatives, chimpanzees [13]. Linguists and cognitive neuroscientists have also determined which regions of the human brain are involved in which parts of speech with some degree of precision. For example, the understanding of proper nouns is thought to occur in the anterior and medial areas of the temporal lobe, and that of common nouns on the lateral and inferior temporal lobes [14].
Thus, language is enabled and constrained by human biology and physiology. The levels of human physiology that enable language stem from the brain and reach to the organs involved in the production of human speech, as well as those for the processing of the speech of others. At a higher level in language semantics than the functional structural underpinning of universal grammar is the language lexicon, which may vary in arbitrary ways from language to language, allowing language history to be traced by shared lexical commonalities. Language can thus be thought of as a complex adaptive system that comprises a series of interacting agents, each of which responds to local conditions following simple rules [15]. As such, language allows people to divide the external world in a myriad of ways and to create artificial worlds, such that it is nearly impossible to imagine a social system comprising human beings that is not ordered by language [16].
While many species on the planet communicate, the cognitive possibilities that spoken and written language affords are thought to be distinctively human. It is unlikely that non‐human animals or indeed any other species has syntactical grammar rules as complex as humans. As Fedurek and Slocome put it: ‘humans can convey an infinite amount of messages using a limited number of words because of the powerful system of grammatical rules that govern the structure and form of language, including the ordering of words into meaningful sentences’ [17]. The anthropologist Leach argues that the transmission of culture, in the sense of patterns of learned behaviour, from generation to generation by learning instead of by genetics, is a characteristic of many other species: ‘bees and ants have extremely elaborate, highly organized social systems which appear to get along without any concept‐forming medium of communication which is even remotely similar to that of human language’ [16]. Thus, it is clear that non‐human languages exist, and that language itself is necessary but not sufficient property of human culture or indeed of animal culture.
How Language Is Transformed by Biology and Physiology

Hauser et al. argue that language as a biological faculty may be divided in ‘Faculty of Language in a Broad Sense’ (FLB) and ‘Faculty of Language in a Narrow Sense’ (FLN) [18]: the former may be analogous to animal communication and include aspects of language faculty (includes the perceptual‐articulatory apparatus); and the latter may be analogous to the human language faculty (and includes the conceptual and intentional apparatus). Kirby [5] and Corballis also assert that recursion is a defining property of human language. Recursion is defined as a procedure in which one of the steps involves invoking the procedure itself, such as that involved in nesting syntax meanings within sentences [19]. However, others hold a view that a reliance on recursion as the main feature of human linguistic difference may oversimplify syntactical aspects of human language, in which communicating propositional structures is an adaptation consisting of many different interacting sub‐systems within human consciousness and physiology. Thus, if there are common features between FLB sense and FLN, then the process of complex language use involving recursion, intentionality, empathy and meta‐representation is a gradual one involving intermediate stages that also have evolutionary correlates and limitations. After all, the most distinctive property of language is that it is generative, allowing parts of language syntax to be recombined to make other language structures, which involves the ‘embedding and concatenation of phrases’ [20]. Consequently, is language use, like evolutionary behaviour, a process of constant adaptation? Is there a plausible evolutionary trajectory from protolanguage to ways of communicating that have yet to be imagined?
As Richardson and Boyd suggest, the division of labour between genes and culture is a co‐evolutionary process [21]. The Baldwin effect argues that language adaptation occurring within an individual’s lifetime may affect linguistic competence: in other words, if language use and linguistic competency give an advantage, a trait change within an organism’s lifetime as a part of cultural evolution can be assimilated into the epigenetic repertoire. As Richardson and Boyd explain, ‘so long as a higher capacity cultural communication system was favoured, cognitive modifications to more efficiently acquire explicitly linguistic features like symbolic words and grammar would be favoured as the system passed some threshold of complexity’ [22]. Thus, it is possible that a Baldwinian effect can lead to random selection for linguistic competencies if they provide an adaptive advantage. In one sense, then, biology and social evolution act as a fixed‐point attractor for human linguistic competency [23]. However, as Yamauchi and Hashimoto point out, it is possible that a strong assimilatory process might be available within 10–20 generations with a selective environment that is dynamically niche‐constructed, but a functional redundancy may result on the genetic information of the learned trait [24]. It is possible that the redundant genetic information becomes degraded while no effect is observed on a linguistic level. For example, the gene for birdsong results in natural selection of related adaptive physiological changes such that a functional redundancy occurs without externally motivated environment changes [25]. Arguably, the exciting things about language use and birdsong might happen in just a cycle of gene recession as other genetic factors are selected. For example, syntactic recursion may facilitate more complex language use that results in a gradual redundancy of selection but an increase in a selection of genes for other language features such as intentionality, with corresponding physiological adaptation in the neural networks.
If language is a form of cultural evolution, it is useful to look at the relationship between its historical progression and changes in human evolutionary physiology. Whilst some observations have been made (such as the descended larynx that enables human speech), it is difficult to isolate any one physiological feature in cognate language use (red deer and lions, for example, also have a descended larynx). More probably, language production is enabled because of the interaction of a number of different musculo‐skeletal and neural features of human physiology, some of which are shared by other species. Thus, language could evolve from a rudimentary set of culturally transmitted signals, impoverished vocalisations or manual gestures to symbolic words and grammar via selective genetic adaptation, given the relative social utility of such communication. However, people are the product of their genes, gene expression and also the environment—all three are involved in the evolutionary process. Cultural evolution leads to the adaptation of a universal grammar. We can ask then the fundamental questions: What is the reason for linguistic diversity? What is the rate of linguistic evolutionary change? How quickly do languages adapt? What is the approximate rate of language change? If these questions we can then begin to understand not only how ‘fixed’ cultural meaning are temporally mutable and may change in time, and also how soon current linguistics constructs may become less intelligible to our ‘future selves’.
Richardson and Boyd suggest that linguistic diversity arises not just because of ‘cultural containment’ by ethnicity or geography, but also for the sociological reason that it limits communication between people who cannot freely trust one another or in which truthful communications lead to maladaptive behaviours [21]. However, this is increasingly tested by conditions of globalisation, in which it could be argued that technology precedes the use of language frequency and adoption of new linguistic constructs. In this way, humans are also different from some other animal species in depending on social transmission for adaptive behaviours.
There is a three‐cycle process of language change [26]. Main determinants are learning (ontogeny), learning produces cultural evolution, (glossogeny) take place with much slower biological evolution and development of phenotype (phylogeny). There are many more changes from language change (ontogeny) and learning at cultural level before physical genetic adaptation and change to phenotype (phylogeny). So both innate endowment and cultural evolution (development of memes, forms of cultural encoding such as library holdings, the development of the lexicon and public memory, etc.) effect language change. Occurs through millennia and through changes at the individual level, the wider cultural level of the (lexicon), and biologically to the species. But ultimately genes encode the linguistic ability of people within the society, although language changes the fitness of the landscape for cultural formations.
Language Adaptation

Intentional agency enters into most human communications. Because of the innate complexity of language, individuals may not have enough in common to facilitate open and honest communication all the time. Furthermore, individuals and societies are competitive and consequently many people have mixed motives in communication, including self‐interest. If not all communication, speech and language is trustworthy, then some features of language may not have a selected advantage for evolution, depending on the cognitive and social cost of information exchange. This leads to the two features of human language use. Firstly, language variation may be adaptive; and secondly, because combinatory communications systems have rules of interpretation, they may be vulnerable to both misunderstanding ‘noise’ and also to ‘deception’. Thus, it requires concerted forms of co‐operation to enable language evolution. Nevertheless, language diversity may not have a basis in issues of trust. Presumably niche‐adaptation is the most likely cause of such diversity, not deviant inventiveness. However, human biological complexity and conscious cognition in language occur on related but different levels of affect.
Coeval with language use, people have the ability to represent different referential locations of time and space through symbolic communication. While the basis of that ability might lie in gene acquisition and physiological capability, the sociality of language is largely separate from biological substrates. Hence, we can imagine that language use evolves in an individual’s lifetime through acquisition of different linguistic competencies: for example, grammar and lexicon (tens of years). This stage is known as ontogeny, possibly leading to genetic adaptation through the Baldwin effect. However, languages evolve over hundreds of years (think, for example, of the differences between Chaucer, Shakespeare and the modern novel). This stage is known as glossogeny. The ability of species to engage with language abilities (such as a universal grammar) is acquired over thousands of years. That is known as phylogeny. MacWhinney adds an epigenetic stage (a combination of environmental and genetic interactions), a developmental (ontogenetic) stage, a processing time‐frame, a social emergence time‐frame and a diachronic (glossogentic) period of language change [27].
However, as Dediu suggests, ‘it is becoming increasingly clear that not only are the pathways connecting genes to phenotypes non‐linear and difficult to map, and that gene‐gene and gene‐environment interactions are the norm in the development of most phenotypic aspects, but also that there is no clear‐cut difference between ‘genetic’ and ‘environmental’, that ‘development’ is not a discrete, encapsulated, and teleological phase in the life cycle of an organism, and that ‘genes’ are essential to all processes at all times’ [12]. It is difficult to tell from evolutionary archaeology exactly how changes in human phylogeny resulted in the genetic selection for complex language ability amongst humans. Fitch suggests that it occurred somewhere between 200,000 years ago and the present day [28]. In most people, the control of language is co‐ordinated by the left brain hemisphere—the regions of the brain which activate and integrate motor control of the lips, tongue and vocal cords. It is the claim of many linguists that human beings are born with a knowledge of linguistic structure and the ability to acquire language (known as a Universal Grammar), which is encoded in the genome at least in part by the FOXP2 gene, which is linked to the linguistic features of the brain found in Broca’s and Wernicke’s areas and the pre‐frontal cortex. The proponents of Universal Grammar (UG) argue that people are born with linguistic competency which has a genetic substrate. Thus, relatively small changes at the phenotypic level can lead to quite large cognitive and behavioural changes. Comparison with other animal species reveals that some animals (such as dolphins and parrots) have quite sophisticated imitative skills, which may imply a form of language‐related empathy; similarly, animals may have ‘functionally referential’ alarm calls capable of bearing external referents. Apes are also capable of audience effects that can bear ‘conspecific’ information. However, as Fitch suggests, many such calls are not intentionally referential and might not shape calling in ways relevant to the listener’s knowledge [29]. This does not necessarily imply that animals cannot individuate at a mental level in their communication, but rather they may not have theories of mind that are capable of attributing more than physiological awareness of others. Thus many animals lack an ability to ‘shape’ their communication in ways that humans do.
For Biological evolution, a canonical interactor is an organism as it interacts with environment, the ecosystem and fellow organisms, in such a way as it causes differential replication [30]. Either organism survives and reproduces or does not survive or reproduce. If it does reproduce, then its genes are differentially replicated. Similarly, in the process of language change, a speaker is interactor, who interacts with environment by virtue of speech and or written communication and whom causes some replication of linguistic variants and not others, resulting language constancy and/or a change [30]. There is (despite the conscious effort and motivation of a speaker and writer) a degree of randomness in replication process and random fluctuations in replicator frequencies. If a fluctuation is greater than zero, replication can take place and if the fluctuation happens to zero, then the replicator extinguished. The process of genetic drift is so termed because that change takes place in population simply due to random processes, and no selection takes place [31].
According to the argument of language evolution as a gradual adaption in human society from genetic assimilation and culturalization, the underlying genetic substrate of language enables a Universal Grammar, which is a set of grammatical principles that applies across all human languages. This goes some way to explaining language diversity—each society develops its own adaptation to a different linguistic environment that fits its locality and social identity. However, language might be shaped by physiological limitations that are not entirely dedicated to language. These include perceptuo‐motor factors, including physiological features that account for the seriality of vocal structures, and perceptual systems that store sensory information (involving the retina, primary visual cortex, occipital lobe, the dorsal stream and the ventral stream; the vocal tract affected by the tongue, whose muscles are controlled by the hypoglossal nucleus and nucleus ambiguous); and lower jaw (controlled by the trigeminal nucleus; and lips controlled by the facial motor nucleus). Second, language might be shaped by cognitive limitations on learning and processing, including memory, and from structures of mental representation and reasoning (involving the cerebellum, lateral hemispheres, Broca’s and Wernicke’s areas, the arcuate fasciculus, and pre‐frontal cortex) [32].
In order to describe the two inter‐related issues of language development, Chater and Christiansen have specified the terms ‘N‐induction’ and ‘C‐induction’. ‘N‐induction’ describes the language challenge of understanding and manipulating representations and relationships in the natural world, and ‘C‐induction’ describes how people acquire ability to converse and co‐ordinate with each other. They make the point that in N‐induction, the world imposes an external standard, but in C‐induction the standard is social. The difference is that in the latter language conditions people to do the same thing but not necessarily the objectively true thing. While both conditions are predicated on learning, the second does not rely only on the correspondence theory of truth but also on ‘relational’ or performativity aspects of language. Chater and Christiansen make the point that language is mostly acquired by C‐induction, given that there is no human‐independent true language which people learn [33]. Rather, they learn the language of their developmental environment (although second‐language learners may have more than one such environment).
Thus in the languages adaptionists’ model, language is guided by genetic inheritance and the growth of cognitive structures (phylogeny), which are shaped by the environment as the learner acquires the language of their caregivers and culture, allowing for language evolution at the level of glossogeny. Such language acquisition contributes to the reproductive potential of the individual. At the level of individual and group language use, language may be seen as a mapping device between meanings and signals. In this way, languages may be composed with a shared signal structure, or be seen as being holistic, whereby such structure may or may not be implicated in a shared meaning space [34].
Language Change
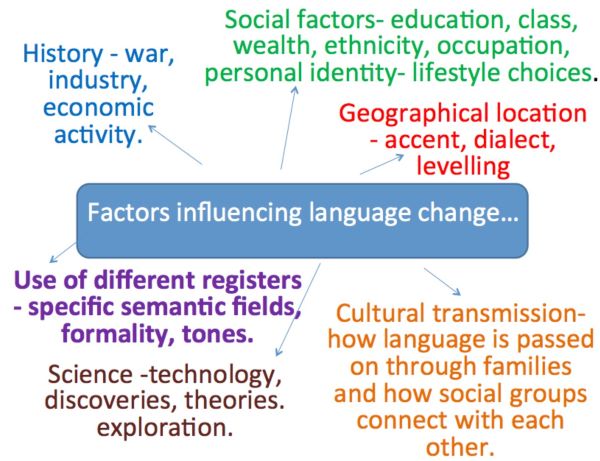
As Kuun points out, acculturation can be a social mechanism that acts as a catalyst for new identities and for language change [35]. When people are speaking the same language they do not necessarily share the same identity. The constructivist approach argues that ethnic identity can change constantly and that everyone identity is subject to change through language [36]. Language can act as a mediator of personal and social identity and where one language is used, multiple identities and second languages may provide a background lexicon to it. Identity is created from language to adapt to different situations. As Kuun argues, language shift process can be influenced by a variety of factors including: immigration, the value of a language and the status of a language for a given community, and also language shift may be determined by the education opportunities of the next generation [37]. Language shift may also be affected by opportunities to speak the same language, the influence of media on language and the opportunities to learn languages. Communities of language use are not homogeneous, there are segments who want to use the existing language, and those who want to change, in‐group or out‐group, strengthening or weakening of identity can result, and traits of identity, and language acquisition and change (e.g. Idiom and dialect) is constructed by language and shared between groups and may be gained and lost or acquired over time [37]. As Stadler et al. suggest ‘… individuals are biased towards variants which have recently seen an increase in their frequency of use’ [38]. Thus, language change may happen relatively quickly in individual terms but its uptake is nevertheless dependent on distribution within a population, which is subject to other temporal pressures on language use. These pressures might include: media use and availability, immigration, inter‐mixing, access to public facility such as education and the extent to which the cultural adoption of language change takes place in a low or high context communication culture.
Emergence of language idiom is determined in part by occurrence of particular variants and frequency of interactions between different speakers [39]. New dialect formations are acquired or lost on the basis of frequency of use, in terms of exposure to tokens of language used by speakers with whom another speaker reacts. The primary or sole factor in new dialect formation is accommodation into language use [40]. Trudgill’s theory is related to the fact that linguistic behaviour is determined by language use in communication interaction [41]. It shares compatibility with neural evolutionary theory, but also implies that propagation of language change is result of the relationship between speakers and social factors [42]. There are debates about what social factors are but most agree that they may include: social class, prestige, group identity and how speakers relate and adopt some linguistic markers but not others. However, according to Trudgill’s theory, frequency of use and accommodation are the sole mechanisms, which account for new dialects evolving in language use [40].
Language change is a two‐step process involving the generation of variations (innovations) and the propagation of a variant through a speech community [42]. The process of linguistic behaviour involves the replication of tokens of linguistic structure such as sounds, words and constructions in language use. Speaker acts form utterances which replicate structures heard before, and structures combine in new ways form novel utterances as the variability of language use is exponential [42]. Tokens of linguistic structure are called a ‘lingueme,’ replication altered by prior replications; linguemes may include phoneme, a morpheme or a whole phrase or any basic unit of a linguistic structure. The replication of one lingueme variant over another is influenced by a number of factors, for example, the rate of exposure to speakers, or social values attached to a lingueme. Hall argues that a second entity or interactor is requires for selection to operate, interactor causal ‘differential replication of replicators’ [43]. Types of interactor selection, some interlocutors will be preferred or dispreferred by the speaker no matter how frequently or infrequently he or she interacts with them. Consequently, some linguistic replications, some utterances are weighted accordingly, and some variants that are weighted more heavily will be differentially replicated [44]. The weighing of existing tokens is relative to new tokens and a reduction in weight of existing tokens, continues every time a new token is added to the speaker’s memory [45]. The rate of decay is controlled by A, and A is small. Interestingly, the resulting decay function is close to the exponential form that is used in psychological models of memory [45].
Entrenchment in memory is dependent on continuously hearing new tokens that are broadly compatible three results. First, exposure to tokens distributed over time rather massed at once facilitates consolidation of memory. Second, a lower limit to the rate of decay is imposed by the consolidation of memory, discussed in terms of a ‘time window’ of lying down of memory. If the time between two tokens of same variant is longer than others with whom they compete for the same ‘time window’, then those tokens do not help consolidate a memory trace [46]. Third, it is the result of linguistic generalization that linguistic forms of a higher token frequency are likely to be retained, whereas linguistic forms with lower token frequency more likely to be replaced by model variants, and low frequency forms in the morphological paradigm. In lower frequency, there is a larger gap between instances of memory formation allowing a greater time for decay between the arrival of individual instances, making it less likely that new tokens contribute to entrenchment in memory [46]. As Moder points out, although chunks may be fixed in form and meaning through a specific context, if component parts maintain their analysability, then the chunk is not immutable. When supported by possible contextual based inferences, the analysability of components (or the parts of a chunk) leads to significant shifts both in meaning of the chunk of compositionality. A strong association of communication with an alternative niche may lead to wide scale shift in form and meaning [47], hence, explaining language change.
Language change can occur relatively quickly—anywhere from decades to centuries, as the meaning of particular phrases morphs in the context of language use (ontogenic changes. For example, Moder points out how the meaning of the phrase ‘begging the question’ has changed over the course of 500 years, with changes in cultural context of language use. The original meaning meant ‘asserting the question’, rather than arguing the merits of position [48] with independent evidence meaning shifted since sixteenth century to include ‘introducing a new question’. Diachronic change can be the result of ‘chunking’ in which formulaic expressions are thought to evolve out of ‘form‐meaning pairings that co‐occur within communicative contexts’ [49] and compositionality, which is the ‘transparency of the meaning of the whole word or phrase … whether or not meaning predicable from the sum of meanings of its component parts’. Language change can come about through loss of chunking and compositionality [50]. However, the underlying evolutional physical structure of language changes very slowly, at only one gene variation in 5000–1000 years (phylogenic change).
As Clark [51] notes, Traugott [52] discusses the role that pragmatics can play in accounts of language change. Prominent are ‘bridging contexts’ in which an utterance can be understood by a listener as having innovative meaning, as well as an earlier meaning. In some linguistic contexts, the innovative meaning may be favoured but remain cancellable and not conventionalised. Some bridging contexts occur when an expression gives rise to a ‘pragmatic implicature’ frequently enough so that the context of implicature becomes associated with the expression. It becomes imprinted to ‘cancel’ the implicature so it is no longer clear that its activation is not a part of an emergent coded meaning. Blakemore distinguishes between a conceptual and procedural meaning [53]. A grammaticalisation, which is a process of language change in which nouns and verbs transform into prepositions and affixes, emerges when there is an addition of procedural information to the semantic of an expression, as well as already encoded information’ [51].
As Operstean notes the main way language changes is through contact based theory as explained in the context of dialect acquisition or change: ‘A language (or dialect) Y at a given time is said to be descended from language (or dialect X) of an earlier time if and only if X developed into Y by an unbroken sequence of instances of native‐language acquisition by children’ [54]. But there are some linguistic variable and language‐transmission situations that don’t fit that strict definition. The acquisition of new dialects, or immigrant koines, for example, begins with mixing, or creation of pool of linguistic features drawn from language varieties present in environment in which language spoken. The mixing stage is followed by levelling, or loss of demographically minority variants [55]. In the new dialect formation, features that get levelled out are those that are”, features that are selected reflect demographic ratios in contact environment, as well as level of constancy such as markedness, perpetual salience, semantic transparency, feature selection guided by ‘founder principle’, in which the linguistic features of founding population may have an advantage in selection [55]. The levelling stage is followed by simplification, or the elimination of marked variants and irregular patterns. The elimination of marked phonemes occurs in phonology (the system of contrasting relationships among speech sounds) and that of marked patterns in morphology (structures of words and parts of words; word relationships). Some of this process may be driven by adult second language acquisition. Some changes might occur from mistakes through slang uptake, others from shifts in meaning context. Formation contact varieties include hybrid forms, inter‐dialect forms, variant forms from original language use which is ‘left over’ after levelling reallocated, pressed into service as new stylist marker, new variety stabilised acquires own forms [56].
Research indicates alternations between periods of abrupt and gradual change correlated with language transmission, intergenerational transmission to young learners during periods of gradual changes a non‐generational transmission to adult learners during periods of rapid change. Operstean hypothesises an alternation between periods of punctuation (new language formation) and equilibrium (gradual compenetration among neighbouring languages) [57].
Intergenerational transmission is dominant mode during periods of gradual change. Gradual compenetration among neighbouring languages takes place through the bi and multi‐lingual individuals, acquired languages as children, happens during stable periods of predominantly generational transmission. Social stability prevailing encourages maintenance of existing norms, and results in slow change. This is contrasted with the punctuation stage, in which the dominant mode is non‐generational, and the breakdown of social structures of events which sets off punctuation brings about the breakdown of linguistic norms. Operstean notes that established language families may be made up of mixture of conservative and innovating languages [58].
Endnotes
- Stadler, K., Blythe, R. A., Smith, K. and Kirby, S. (2016). ‘Momentum in Language Change.’ A Model of Self‐Actuating S‐shaped Curves. Language Dynamics and Change, 6: 172.
- Baronchelli, A., Chater, N., Pastor‐Satorras, R. and Christiansen, M. H. (2012). ‘The Biological Origin of Linguistic Diversity.’ Plos One, 7 (10): 1.
- Fedurek, P. and Slocome, K. E. (2011). ‘Primate Vocal Communication: A Useful Tool for Understanding Human Speech and Language Evolution?’ Human Biology, 83 (2): 271.
- Jakobson, R. (1970). ‘La Linguistique.’ In Tendances Principales De La Recherché Dans Les Sciences Sociales et Humanes. Paris: UNESCO: 530.
- Kirby, S. (2007). ‘The Evolution of Language.’ In Oxford Handbook of Evolutionary Psychology, edited by R. Dunbar, and L. Barrett, 669-81. Oxford: Oxford University Press.
- Smith, K., Brighton, H. and Kirby, S. (2003). ‘Complex Systems in Language Evolution: The Cultural Emergence of Compositional Structure.’ Advances in Complex Systems, 6 (4): 537.
- Fedurek, P. and Slocome, K. E. (2011). ‘Primate Vocal Communication: A Useful Tool for Understanding Human Speech and Language Evolution?’ Human Biology, 83 (2): 153.
- Smith, K., Brighton, H. and Kirby, S. (2003). ‘Complex Systems in Language Evolution: The Cultural Emergence of Compositional Structure.’ Advances in Complex Systems, 6 (4): 538.
- Starte, L. (2006). ‘Echoes and Reflections.’ On Media Ecology as a Field of Study. Cresskill, NJ: Hampton Press Communication Series: 45.
- Starte, L. (2006). ‘Echoes and Reflections.’ On Media Ecology as a Field of Study. Cresskill, NJ: Hampton Press Communication Series: 46.
- Berwick, R. and Chomsky, N. (2011). ‘The Biolinguistic Program: The Current State of Its Development.’ In The Biolinguistic Enterprise: New Perspectives on the Evolution and Nature of the Human Language Faculty, edited by A‐M. Di Sciullo, and C. Boeck, 19-41. Oxford: Oxford University Press: 20-21.
- Dediu, D. 2011. ‘Are Languages Really Independent From Genes? If Not, What Would a Genetic Bias Affecting Language Diversity Look Like?’ Human Biology, 83 (2): 281.
- Schoenemann, T. P. (2009). ‘Evolution of Brain and Language.’ Language Learning, 59 (1): 175.
- Schoenemann, T. P. (2009). ‘Evolution of Brain and Language.’ Language Learning, 59 (1): 176.
- Schoenemann, T. P. (2009). ‘Evolution of Brain and Language.’ Language Learning, 59 (1): 163.
- Leach, E. (1982). ‘Social Anthropology.’ New York: Oxford University: 107.
- Fedurek, P. and Slocome, K. E. (2011). ‘Primate Vocal Communication: A Useful Tool for Understanding Human Speech and Language Evolution?’ Human Biology, 83 (2): 166.
- Hauser, M. D., Chomsky, N. and Fitch, T. W. (2002). ‘The Faculty of Language: What Is It, Who Has It, and How Did It Evolve?’ Science 298 (22): 1569.
- Corballis, M. C. (2009). ‘The Evolution of Language.’ The Year in Cognitive Neuroscience: Annals of the New York Academy of Science, 1156: 36.
- Corballis, M. C. (2009). ‘The Evolution of Language.’ The Year in Cognitive Neuroscience: Annals of the New York Academy of Science, 1156: 36.
- Richardson, P. J. and Boyd, R. (2010). ‘Why Possibly Language Evolved.’ Biolinguistics, 4 (2-3): 299.
- Richardson, P. J. and Boyd, R. (2010). ‘Why Possibly Language Evolved.’ Biolinguistics, 4 (2-3): 290.
- Johnson, N. (2012). ‘Simply Complexity.’ A Clear Guide To Complexity Theory. London: One World Publications: 4.
- Yamauchi, H. and Hashimoto, T. (2010). ‘Relaxation of Selection, Niche Construction, and The Baldwin Effect of Language Evolution.’ Artificial Life, 16 (4): 271-287.
- Yamauchi, H. and Hashimoto, T. (2010). ‘Relaxation of Selection, Niche C, and The Baldwin Effect of Language Evolution.’ Artificial Life, 16 (4): 284.
- Smith, K., Brighton, H. and Kirby, S. (2003). ‘Complex Systems in Language Evolution: The Cultural Emergence of Compositional Structure.’ Advances in Complex Systems, 6 (4): 541.
- MacWhinney, B. (2005). ‘The Emergence of Linguistic Form in Time.’ Connection Science, 17 (3-4): 191-211.
- Fitch, W. T. 2010. ‘The Evolution of Language.’ New Scientist Instant Expert I: viii. http://www.newscientist.com/data/doc/article/dn19554/instant_expert_6_‐_the_evolution_of_language.pdf.: 203.
- Fitch, W. T.. 2010. ‘The Evolution of Language.’ New Scientist Instant Expert I: viii. http://www.newscientist.com/data/doc/article/dn19554/instant_expert_6_‐_the_evolution_of_language.pdf.: 205.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of the Emergence of New Zealand English.’ Language Variation and Change, 21: 269.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of the Emergence of New Zealand English.’ Language Variation and Change, 21: 270.
- Chater, N. and Christiansen, M. H. 2009. ‘Language Acquisition Meets Language Evolution.’ Cognitive Science, 34: 1135-6.
- Chater, N. and Christiansen, M. H. 2009. ‘Language Acquisition Meets Language Evolution.’ Cognitive Science, 34: 1141.
- Smith, K., Brighton, H. and Kirby, S. (2003). ‘Complex Systems in Language Evolution: The Cultural Emergence of Compositional Structure.’ Advances in Complex Systems, 6 (4): 544.
- Kuun, E. (2015). ‘Impact of Change of The Language Environment on The Extent of Language Shift in The Context of Linguistics.’ TRAMES, 19 (69/64), 1: 73.
- Kuun, E. (2015). ‘Impact of Change of The Language Environment on The Extent of Language Shift in The Context of Linguistics.’ TRAMES, 19 (69/64), 1: 74.
- Kuun, E. (2015). ‘Impact of Change of the language Environment on the Extent of language Shift in the Context of Linguistics.’ TRAMES, 19 (69/64), 1: 76.
- Stadler, K., Blythe, R. A., Smith, K. and Kirby, S. (2016). ‘Momentum in Language Change.’ A Model of Self‐Actuating S‐shaped Curves. Language Dynamics and Change, 6: 192.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of The Emergence of New Zealand English.’ Language Variation and Change, 21: 257.
- Trudgill, P. (2004). ‘New‐dialect Formation,’ The Inevitability of Colonial Englishes. Edinburgh: Edinburgh University Press.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of The Emergence of New Zealand English.’ Language Variation and Change, 21: 258.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of The Emergence of New Zealand English.’ Language Variation and Change, 21: 268.
- Hall, R. (1958). ‘Creolized Languages and ‘Genetic Relationship.’ Word, 14: 269.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of The Emergence of New Zealand English.’ Language Variation and Change, 21: 271.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of The Emergence of New Zealand English.’ Language Variation and Change, 21: 275.
- Baxter, G. J., Blythe, R. A., Croft, W. and McKane, A. J. (2009). ‘Modeling Language Change: An Evaluation of Trudgill’s Theory of The Emergence of New Zealand English.’ Language Variation and Change, 21: 257-296.
- Moder, C. L. (2016). ‘Begging the Question: Chunking, Compositionality and Language Change.’ European Journal of English Studies, 20 (1): 45.
- Moder, C. L. (2016). ‘Begging the Question: Chunking, Compositionality and Language Change.’ European Journal of English Studies, 20 (1): 36.
- Moder, C. L. (2016). ‘Begging the Question: Chunking, Compositionality and Language Change.’ European Journal of English Studies 20 (1): 37.
- Moder, C. L. (2016). ‘Begging the Question: Chunking, Compositionality and Language Change.’ European Journal of English Studies 20 (1): 42.
- Clark, B. (2015). ‘Relevance Theory and Language Change.’ Lingua, 143: 175-176.
- Traugott, E. C. (2012). Pragmatics and language change. In Allen, K., Jaszczolt, K. (Eds.) Cambridge Handbook of Pragmatics. Cambridge: Cambridge University Press.
- Blakemore, D. (2007). Constraints, concepts and procedural encoding. In Burton‐Roberts, N. (Ed.). Pragmatics. Basingstoke: Palgrave Macmillan: 45-66.
- Operstean, N. (2014). ‘Contact‐Genetic Linguistics: Toward a Contact‐Based Theory of Language Change.’ Language Sciences, 48: 2.
- Operstean, N. (2014). Contact‐Genetic Linguistics: Toward a Contact‐Based Theory of Language Change.’ Language Sciences, 48: 4.
- Operstean, N. (2014). ‘Contact‐Genetic Linguistics: Toward a Contact‐Based Theory of Language Change.’ Language Sciences, 48: 5.
- Operstean, N. (2014). ‘Contact‐Genetic Linguistics: Toward a Contact‐Based Theory of Language Change.’ Language Sciences, 48: 1-15.
- Operstean, N. (2014). ‘Contact‐Genetic Linguistics: Toward a Contact‐Based Theory of Language Change.’ Language Sciences, 48: 10.
Chapter 2 from Sociolinguistics: Interdisciplinary Perspectives, originally published by IntechOpen, 07.05.2017, under the terms of a Creative Commons Attribution 3.0 Unported license.






